Scgf gene expression is down-regulated in the tumorigenic canine osteosarcoma cells relative to their non-tumorigenic cells (549). Scgf-highly positive MSC subpopulation is predominantly present in human osteoblastic osteosarcoma tissues (638). Malignant pleural effusion secondary to osteosarcoma contains less tumor-associated scgf+ B cells and macrophages than the primary tumor tissue (682). When GD2+ U2OS osteosarcoma cells are cocultured for 3 days with combination of N-803 (IL-15N72D mutein superagonist coupled with IL-15RαSu/IgG1Fc), anti-GD2 MoAb dinutuximab and ex vivo expanded NK cells, NK cell-mediated cytotoxicity is enhanced against U2OS cells to reduce their SCGF secretion (656).
Wnt (wingless & int-related protein) gene has a commitment to the fate of stem cells in fetal tissues. Of wnt, wnt5a gene is specifically up-regulated in the osteoblasts differentiated from MSCs. Scgf gene is down-regulated in the calvaria of wnt5a-/- mice as compared with that of wnt5a+/+ mice (43). SCGF closely interacts with Wnt5a in osseous differentiation.
Administration of HDAC inhibitor, Entinostat/MS-275, in utero up-regulates scgf gene expression in calvaria from wild type and Runx2+/- cleidocranial dysplasia model mice (557).
SCGF level in the plasma and synovial fluid from patients with knee osteoarthritis (OA) or rheumatoid arthritis (RA) is eight times higher than that of healthy controls (360). Synovial epithelial cells from RA patients exhibit higher expression of SCGF than those from OA patients (Pat32). Higher level of SCGF in the synovial fluid from knee OA patients correlates significantly with OA severity, knee pain and function (605). Male OA patients have a higher knee synovial fluid SCGF level than female patients (523). Scgf gene expression is up-regulated in the synovial tissues from pristane-induced severe erosive RA model DA rats compared to mild non-erosive DA.F344(Cia5a) rat controls (399).
Interlaminar epidural steroid injection significantly ameliorates low back pain and lowers plasma SCGF level 7 to 10 days after the therapy for patients with intervertebral disc disorders, particularly disc herniation (513). Scgf over-
expression is associated with intervertebral disc degeneration since plasma SCGF level is elevated in intervertebral disc herniation and scgf knockdown with scgf siRNA down-regulates several inflammatory mediators in human nucleus pulposus cells (709).
Bone-invasive type of clivus chordoma up-regulates SCGF production relative to non-invasive type (494).
Scgf gene is expressed in the multinucleated giant cells from benign maxillofacial and axial/appendicular giant cell bone tumors (545).
Scgf gene expression is significantly down-regulated in the ligament tissues from patients with chronic phase of anterior cruciate ligament tear compared to acute phase (582).
Kidney, Bladder
HLA-matched allogeneic non-ablative PB-SCT and donor lymphocyte transfusion is performed to the patients with stage IV renal cell carcinoma. Graft versus tumor reactive donor CD8+ CTLs are cloned from the patients whose tumor is involuted, and they recognize minor H antigen on chromosome 19q13, particularly KLK and scgf (196). SCGF-driven growth of renal carcinoma cells may be a target for CD8+ CTL. SCGF level in the plasma from patients with metastatic renal cell carcinoma is not altered before and after treatment with sorafenib or sorafenib+interferon-Α compared to normal controls (354).
Scgf gene expression in renal cancer tissues usually indicates unfavorable prognosis (562).
Serum SCGF level 48 hours post-renal transplantation in patients with delayed or slow graft function is significantly higher than those with immediate graft function (286). Plasma SCGF level is significantly elevated within 24 hours after cardiac surgery in patients with the surgery-associated acute kidney injury in proportion to injury stages and serum creatinine levels (633).
Scgf gene expression is highly down-regulated in the skin tissues from chronic kidney disease model mice fed with high phosphate diet, and the dermal scgf down-regulation is further accelerated in the FGF-23 knockout mice fed with high phosphate diet (581).
Scgf gene expression is up-regulatedin vitro in rainbow trout head kidney cells in response to fungal PAMPs
including β-glucan peptide and furfurman (663). SCGF could exhibit innate immunity against fungus infection, e.g.
Malassezia furfur, like same C-type lectin dectin-2 that is a receptor for furfurman.
Scgf gene is up-regulated in the rat bladder cancer induced with 4-OH-butyl (butyl) nitrosamine (158) and human bladder cancer tissue (305). GEO data set GSE31684 shows that scgf up-regulation is associated with bladder cancer progression and metastasis (505).
SCGF level is significantly elevated in the bladder specimens from patients with painful bladder syndrome/interstitial cystitis relative to normal controls (422).
Neuroblastoma
Scgf gene expression is not up-regulated in INSS (international neuroblastoma staging system) stage 1 but in stage 3 and 4 neuroblastoma, indicating that scgf can be MAF (metastasis associated fibroblasts) gene signature in neuroblatoma (287, GEO DataSets GSE3960).
Scgf gene expression is down-regulated in SH-SY5Y human neuroblastoma cells treated with aluminum-Β1-42-amyloid complex but not with aluminum or Β1-42-amyloid (307).
Histone deacetylase inhibitor, trichostatin A (TSA), up-regulates scgf gene expression in BE(2)-C neuroblastoma cells (283).
Gastrointestinal tract
Scgf gene is up-regulated in the biopsy specimen of colon cancer (165), but down-regulated in the 5-FU resistant colon cancer cell line, CCL-227 (163). The latter finding is in conflict with scgf up-regulation in the drug surviving cells (151). Oncogene is often expanded near the fragile site of cancer cell chromosome. Scgf gene is little altered in the vicinity of dhfr-inserted fragile site of MTX-resistant human rectal cancer cell line, HCT116+chr3, randomly inserted with dhfr (197).
No unique somatic mutation of scgf gene has been reported in colon adenocarcinoma (Wellcome Trust Sanger
Institute).
Primary colorectal cancer (CRC) tissues but not adjacent normal tissues produce a high level of SCGF (466). SCGF is produced and secreted by human CRC cell line Colo205 (460), SW620 (189, 255) and RKO (289). SCGF interacts with splice variant 2 but not with splice variant 1 of methionine adenosyltransferase 2β up-regulated in colon cancer cell line, RKO (257), which could regulate transcription and give a growth advantage to cancer cells. Plasma SCGF level is suppressed to rise after patients with metastatic CRC are treated with chemotherapy plus bevacizumab (anti-VEGFMoAb), but elevate along tumor progression (231). Plasma SCGF level is significantly higher in metastatic CRC patients with high peripheral blood neutrophil-lymphocyte ratio (NLR) >5 than those with low NLR ≤5 (488), and can be a predictor of poor survival for metastatic CRC (509). A subgroup of CRC with a molecular signature of scgf up-regulation is a low proliferative, chemotherapy-resistant mesenchymal type with poor prognosis (431). Scgf is identified as a signature gene of higher risk of relapse and intestinal stem cell nature in CRC (491). Scgf gene expression in CRC tissues usually indicates unfavorable prognosis (562).
Hypoxia-induced HIF1A up-regulates scgf in cancer-associated fibroblasts. Secreted SCGF binds to LGR5 on CRC cells to promote VEGFC secretion through activation of Wnt/β-catenin pathway, and facilitates lymphangiogenesis and lymphatic metastasis (712).
Serum SCGF level is not significantly different between pediatric patients with Crohn's disease or ulcerative colitis and healthy controls (498).
Gastrointestinal stromal tumor (GIST) is characterized by kit gene activation, including wild type without mutation, mutation type of exon 9 or exon 11. Scgf gene is up-regulated in the exon 9-mutated GIST as compared with the exon 11-mutated GIST (164). Protein extracts from imatinib-responsive but not from imatinib-unresponsive and untreated GIST tissue samples contain a large amount of SCGF. SCGF is histochemically detected in the myxohyaline stroma with CD68+ macrophages of imatinib-responsive GIST tissues, but not in CD117+ cellular area of untreated and imatinib-unresponsive GIST tissues. Monocytes/macrophages infiltrate to remove cell debris and secrete SCGF through inflammatory reaction subsequent to imatinib-induced apoptosis of GIST cells (339). Responsiveness of GIST to imatinib therapy parallels an accumulation of SCGF in the GIST tissues following imatinib treatment.
Ag-1hiB220+CD11cint plasmacytoid DCs express scgf gene more abundantly than CD11chiCD103hi conventional DCs in colon-draining mesenteric lymph nodes from Citrobacter rodentium-infected mice (525).
NF-κB activation as seen in API2-MALT1+ gastric MALT lymphoma is linked to down-regulation of scgf gene at chromosome 19q13.2-q13.4 (232).
Methylated scgf can be a discriminant methylome in the tumor tissues and peripheral blood white blood cells from patients with gastric adenocarcinoma (593). Scgf is a signature gene for poor prognosis in gastric cancer (699). When up-regulated scgf is knocked down with scgf siRNA (696) and shRNA (699) in MGC-803, AGS and HGC-27 gastric cancer cells, respectively, their proliferation is suppressed, indicating that scgf is a potential therapeutic target for gastric cancer (696, 699). When scgf shRNA-treated MFC mouse gastric cancer cells are inoculated s.c. into nude mice, tumor growth is markedly suppressed with normalized tumor microenvironment immune cells compared to controls (699). SCGF production by MKN7 and MKN45 gastric cancer cells is highly repressed relative to HFE145 normal gastric epithelial cells (279). This is the only report that SCGF is down-regulated in malignant cells.
SCGF level is significantly higher in the serum from patients with esophageal achalasia than healthy controls (620).
Serum SCGF level is higher in Malawian children with nontyphoidal Salmonella infection as compared to convalescent controls (528).
Scgf gene expression is up-regulated in all 4 subtypes of pancreatic adenocarcinoma, including squamous, pancreatic progenitor, immunogenic and aberrantly differentiated endocrine exocrine types (522). TGF-β-Smad signal transduction is one of the pathways to inhibit tumor growth. TGF-β down-regulates scgf gene expression in the Smad4-inhibited pancreatic cancer cell line, PANC-1 (166), indicating that TGF-β tumor inhibition is Smad4-independent and is rationally synergistic with scgf gene down-regulation. SCGF is significantly down-regulated in the supernatants of pancreatic adenocarcinoma-associated fibroblasts cultured with G protein-coupled somatostatin receptor sst1 agonist, SOM230 (645). Scgf is significantly hypermethylated in secretin-stimulated pancreatic juice from pancreatic cancer patients relative to chronic pancreatitis patients and normal controls (515). High plasma level of CA19-9 in combination with methylated scgf DNA accurately detects advanced stages of pancreatic adenocarcinoma (649). SCGF level in the cystic fluid is indistinguishable between pancreatic branch intraductal papillary mucinous neoplasm and inflammatory cyst (380). Cocultures of pancreatic stellate cells with PSN-1, MiaPaCa-2 or L3.6pl pancreatic cancer cells suppress SCGF production compared to combined each cell monocultures (480).
Serum SCGF level shows no significant difference between mild/moderate and severe acute pancreatitis (457).
Lipotoxic stress, e.g. high fat diet-induced obesity and in vitro treatment with palmitic acid, down- and up-regulates scgf and lipid metabolism-related SREBP-1c, Fasn, Plin2 genes in mouse pancreatic islet stellate and β cells, respectively to suppress their proliferation (609). rh/rmSCGF mitigates the lipotoxicity on islet cells, and further activates ERK, AKT, FoxO1 to stimulate their proliferation. (Illustrated in"Biologic Function"section)
Influenza A virus H1N1 and H3N2-infected human pancreatic islet cells secrete a significantly higher amount of SCGF than mock-infected islets (397).
Scgf gene expression is up-regulated in the penile cavernous endothelial cells from streptozotocin-induced diabetic mice when the cells are exposed to high concentration of glucose, and transfection with ninjurin 1 siRNA normalizes the scgf up-regulation (464).
Mouse Krebs-2 ascites carcinoma cells internalizing dUTP-5'-TAMRA-labeled human 500bp Alu repeat are tumor-initiating stem cells that specifically express "stemness" scgf gene (555).
Prostate
Stat3 but not stat5 regulates scgf gene expression in CWR22Rv1 human prostate cancer cells (245).
Scgf gene expression is differentially up-regulated in the hormone-refractory PC3 prostate cancer cells (460) when treated with 150μM abacavir, a nucleoside reverse transcriptase inhibitor for retrotransposon (293). Scgf gene expression is up-regulated in rat UMR-106 osteoblasts co-cultured with human PC3 osteolytic prostate cancer cells through micropore filter (510).
Scgf gene expression is up-regulated in LNCaP human prostate cancer cells in response to androgen, the up-regulation of which is inhibited by treating them with an indirect androgen receptor cotranscription factor SRF (serum response factor) siRNA (309). The findings indicate that scgf is an SRF-dependent androgen-responsive gene signature in prostate cancer and could be a potential therapeutic target.
SCGF is present in the exosomes from human prostate cancer DU145 (554) and LNCaP (567) cells.
Testis
Scgf gene expression in testicular cancer tissues usually indicates unfavorable prognosis (562).
RT-PCR and histochemical studies demonstrate that testicular seminoma produces SCGF (167). SCGF can be one of germinal cell markers. Elevated levels of plasma SCGF are correlated with worse overall survival in patients with metastatic testicular germinal cell tumor (559).
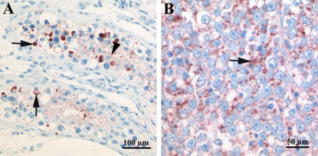
Cytoplasmic SCGF in testicular tumor
Differential methylation is epigenetically found at scgf promoter in the sperm from F3-generation rat after exposure of pregnant F0-generation rat to pesticide (363).
Ovary
Estrogen receptor-positive BG-1 human ovarian adenocarcinoma cells are triggered by 17Β-estradiol to undergo epithelial-mesenchymal transition (EMT), down-regulate E-cadherin gene expression and acquire metastatic potential, when EMT-related genes include scgf up-regulation (225,331). Scgf gene expression is up-regulated in serous and endometrioid ovarian cancer cells (317). Causal regulator active in the tumor tissue from patients with serous ovarian cancer, TNFSF11, up-regulates scgf gene expression (383).
A unique somatic mutation of scgf gene, missense substitution of H281P due to substitution of a842c, has been reported in ovarian serous cystadenocarcinoma (Wellcome Trust Sanger Institute).
Ovarian chorioallantoic membrane cells derived from borderline ovarian tumor express scgf gene (530).
Scgf gene is hypermethylated in the ovary from neonatal rats exposed to high dose of pesticide methoxychrol during fetal and neonatal stages (406).
Uterus, Placenta
Scgf gene is significantly down-regulated when heme oxygenase-1 (HO-1) is knocked down in BeWo choriocarcinoma cells by transduction with a miRNA adapted HO-1-targeting shRNA, relative to HO-1+ control cells with LMP vector (264). Scgf participates with HO-1 in promoting tumor cell growth. Scgf gene expression is apparently down-regulated in tumorigenic HeLa x fibroblast hybrid CGL3 cells and parental HeLa cells compared to non-tumorigenic HeLa x fibroblast hybrid 444 cells (428).
SCGF is produced and secreted by human choriocarcinoma-derived trophoblast JEG-3 cells, but not by SV40-immortalized trophoblast HTR8/SVneo cells (473).
CpG islands within scgf gene are significantly hypomethylated in the endometrial cells at early secretory phase from patients with severe endometriosis relative to healthy controls (535). SCGF level is significantly higher in the peritoneal fluid (621) from infertile patients with endometriosis than those without endometriosis irrespective of disease severity and menstrual cycle stage, indicating that SCGF can be a predictive marker for endometriosis (558). In contrast, SCGF level is significantly lower in the menstrual blood from patients with endometriosis than healthy controls (689).
Scgf gene expression is down-regulated in PB-MNCs from preeclampsia patients at 16 weeks of gestation relative to normotensive controls (302). In contrast, serum SCGF level is significantly higher in patients with preeclampsia than normotensive controls (688).
SCGF level is significantly low in the cervicovaginal fluid with detectable PSA, a marker within 48hrs after sexual intercourse, from women with bacterial vaginosis (653).
Scgf gene is differentially expressed in the umbilical cord depending upon gestational age (382).
Etc.
There have been some sporadic reports concerning scgf gene in diseases.
(1) Batten disease is a neurodegenerative lysosomal storage disorder of lipofuscin due to mutated CLN3 gene. Scgf
gene is down-regulated in the brain of CLN3 knockout mice (198).
(2) Scgf gene is up-regulated in the hyperplastic lymphoid tissue of TNF-α transgenic mice (89).
(3) Scgf is one of overexpressed genes in some cases of desmoid tumor (aggressive fibromatosis) (199).
(4) IL-10 down-regulates scgf gene expression in human PB-MNCs (195).
(5) Hypoxia down-regulates scgf gene expression in the fetal rat intestine (111).
(6) Scgf gene expression is somewhat up-regulated in the muscle samples from patients with Duchenne muscular
dystrophy relative to normal controls (346).
(7) Scgf gene expression is down-regulated in the trabectedin-resistant myxoid liposarcoma 402-91/ET cells relative
to the original sensitive 402-91 cells (366).
(8) Deletion of histone H2B ubiquitin ligase Bre1 by RNAi down-regulates scgf gene expression in mouse fibrosarcoma
RIF-1 cells (367).
(9) Myopathy
Scgf gene expression is kept to be up-regulated in the rat myosatellite cells for 2 to 6 weeks after lacerating
medial gastrocnemius muscle with tibial nerve cut as compared to the lacerated muscle with nerve preserved
intact (371).
Serum SCGF level is significantly higher in MDA5+ autoimmune myositis than healthy controls and ARS+ or
TIF1γ+ cases (674).
(10) Scgf gene expression is up-regulated in the EBV-transformed cells from individuals with 17q21.31 microdeletion
syndrome, of which KANSL1 mutation is causative, compared to normal controls (375).
(11) SCGF level in the serum from patients with mitochondrial diseases is lower than other disease controls (482,
Pat28).